You could make a fascinating study (and many have) just looking at the history of which dinosaurs have held the foreground of popular culture when. The
Iguanodon and
Megalosaurus of the late 1800s, the
Trachodon and
Palaeoscincus of the earlier 1900s, the stratospheric rise of
Velociraptor (
sensu lato) with the release of
Jurassic Park. And then there are those that never quite seem to get their dues. I've
commented before on the odd relegation of
Camarasaurus to the status of also-ran among famous sauropods. But perhaps the ultimate example of a dinosaur forced unfairly to the background is the should-be darling of the Late Triassic,
Plateosaurus.
Plateosaurus 'engelhardti' in the Sauriermuseum at Frick, copyright Ghedoghedo.
Plateosaurus should, by all rights, be a superstar of dinosaur pop-culture. It was one of the first dinosaurs to reach massive size, extending up to nine metres in length and probably standing about as high (or slightly higher) than a tall man at the withers (Yates 2003). It is known from literally hundreds of specimens, many of them with large parts of the skeleton preserved, representing ages from juvenile to full maturity. Some of the bonebeds where it is found contain little but
Plateosaurus and may have been formed in dramatic mass mortality events.
Plateosaurus is easily the best known of the basal Sauropodomorpha, the 'prosauropods'. And yet, though
Plateosaurus regularly appears in popular depictions, it rarely seems to make much more than a brief cameo. Why is this the dinosaur that gets no respect?
In part, it may be because it comes from a time period that gets less attention as a whole. The Triassic tends to get seen as a meer prelude to later, more 'exciting' parts of the Mesozoic.
Plateosaurus itself, together with the other 'prosauropods', tends to also get overshadowed by its later, more eye-catching relatives, the sauropods. And when you get down to it,
Plateosaurus may also be let down by the fact that it is perhaps the single most
average dinosaur you could possibly imagine. Honestly, if you asked someone to depict a truly generic dinosaur, I don't think it would come out looking too different from
Plateosaurus.
Reconstructed Plateosaurus, albeit in a now-obsolescent quadrupedal pose, copyright Elekes Andor.
All these criticisms aside,
Plateosaurus is still a fascinating genus. Its remains have been found across central Europe, in Germany, Switzerland and France. The exact number of species in the genus has long been uncertain. As with other early-named dinosaur genera, 19
th Century palaeontologists named several species whose application has been subject to debate. Yates (2003) recognised two species in the genus, the earlier and smaller
P. gracilis, and a larger, later species that Yates labelled
P. engelhardti but which, due to various taxonomic shenanigans, should probably now be called
P. trossingensis.
Plateosaurus trossingensis is the better known of the two species, known from extensive bone-beds found at Trossingen and Halberstadt in Germany, and Frick in Switzerland (Lallensack
et al. 2021). Some have questioned whether all these bone-beds represent a single species but Lallensack
et al. found that examination of skulls from different locations failed to identify specific distinctions. Both
Plateosaurus species would have been among the largest land animals of their times; even the smaller
P. gracilis may have still reached lengths of five or six metres.
Plateosaurus had a relatively long, narrow head though comparison of this feature with other prosauropods may be complicated by post-mortem distortion.
The life posture of
Plateosaurus has historically been the subject of much dispute, whether it was bipedal, quadrupedal, or shifted freely between the two. However, recent models of the range of movement of the
Plateosaurus hand and fore-arm have concluded that it was incapable of turning its hands palm-downwards, so it could not have supported itself comfortably on its fore limbs (Reiss & Mallison 2014). Obviously, the capacity for quadrupedal locomotion would evolve at some point in sauropodomorph evolution (in this day and age, I don't think anyone is proposing bipedal sauropods) but it was not before
Plateosaurus.
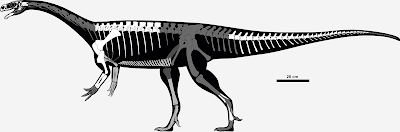
Skeletal reconstruction of Unaysaurus talentinoi, copyright Maurissauro.
The phylogenetic relationships of
Plateosaurus to other sauropods have been similarly disputed.
Plateosaurus is, of course, the type genus of the family Plateosauridae but the concept of that family has varied significantly over time. For a large part of the twentieth century, 'Plateosauridae' was kind of a catch-all for all moderately large prosauropods, with Anchisauridae for the smaller species and Melanorosauridae for the giants. Redefinition of Plateosauridae to include only close relatives of
Plateosaurus have significantly winnowed its contents. The current closest known relative of
Plateosaurus is the recently described
Issi saaneq, based on a pair of near-complete skulls from Greenland (Beccari
et al. 2021). This species is close enough to
Plateosaurus that its remains were previously assigned to
P. englehardti. Offhand, "
issi saaneq" is translated by the species' authors as "cold bone" in the local Kalaallisut language, but this looks to be another situation like "
mei long" where a phrase was converted into a species name without considering that noun and descriptor order is reversed in biological names.
Other likely plateosaurids include two South American species,
Unaysaurus tolentinoi and
Macrocollum itaquii. The status of an Indian species
Jaklapallisaurus asymmetrica is more uncertain. Beyond this, things become increasingly dodgy with little agreement over the details of prosauropod phylogeny. The overall conservative appearance of prosauropods means that phylogenetic studies are heavily reliant on fine details of the osteology that are debated between authors or not preserved in key taxa. Nevertheless, it does appear that the plateosaurids were widespread in the Norian epoch of the Triassic, and are bound to catch the attention of time travellers to the period.
REFERENCES
Beccari, V., O. Mateus, O. Wings, J. Milàn & L. B. Clemmensen. 2021.
Issi saaneq gen. et sp. nov.—a new sauropodomorph dinosaur from the Late Triassic (Norian) of Jameson Land, central east Greenland.
Diversity 13: 561.
Lallensack, J. N., E. M. Teschner, B. Pabst & P. M. Sander. 2021. New skulls of the basal sauropodomorph
Plateosaurus trossingensis from Frick, Switzerland: is there more than one species?
Acta Palaeontologica Polonica 66 (1): 1–28.
Reiss, S., & H. Mallison. 2014. Motion range of the manus of
Plateosaurus engelhardti von Meyer, 1837.
Palaeontologica Electronica 17 (1): 12A.
Yates, A. M. 2003. The species taxonomy of the sauropodomorph dinosaurs from the Löwenstein Formation (Norian, Late Triassic) of Germany.
Palaeontology 46 (2): 317–337.


























{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}